Have you ever wondered if flies can hear you swatting at them? It turns out, they absolutely can! Understanding how flies hear not only reveals fascinating aspects of insect biology but also provides valuable insights into the complexities of hearing in mammals, including humans. Groundbreaking research into the auditory system of the common fruit fly (Drosophila melanogaster) is helping scientists unravel the fundamental mechanisms of hearing and even shed light on human conditions like Usher syndrome, a leading cause of combined deafness and blindness.
The Surprising Similarities Between Fly and Mammalian Hearing
While flies lack external ears like ours, they possess sophisticated auditory organs that function in remarkably similar ways to our own. In mammals, sound detection occurs in specialized “hair cells” within the inner ear. These cells are equipped with tiny, hair-like projections called stereocilia. At the tips of these stereocilia are mechanoelectrical transduction (MET) channels, which are crucial for converting the mechanical energy of sound waves into electrical signals that our brains can interpret.
A group of proteins associated with Usher syndrome type 1 (USH1) is essential for the proper functioning of these MET channels. These USH1 proteins form a complex at the tips of stereocilia and are vital for maintaining the structural integrity of these delicate structures. When sound waves deflect stereocilia bundles, it stretches them, opening the MET channels and triggering the electrical signals that allow us to hear.
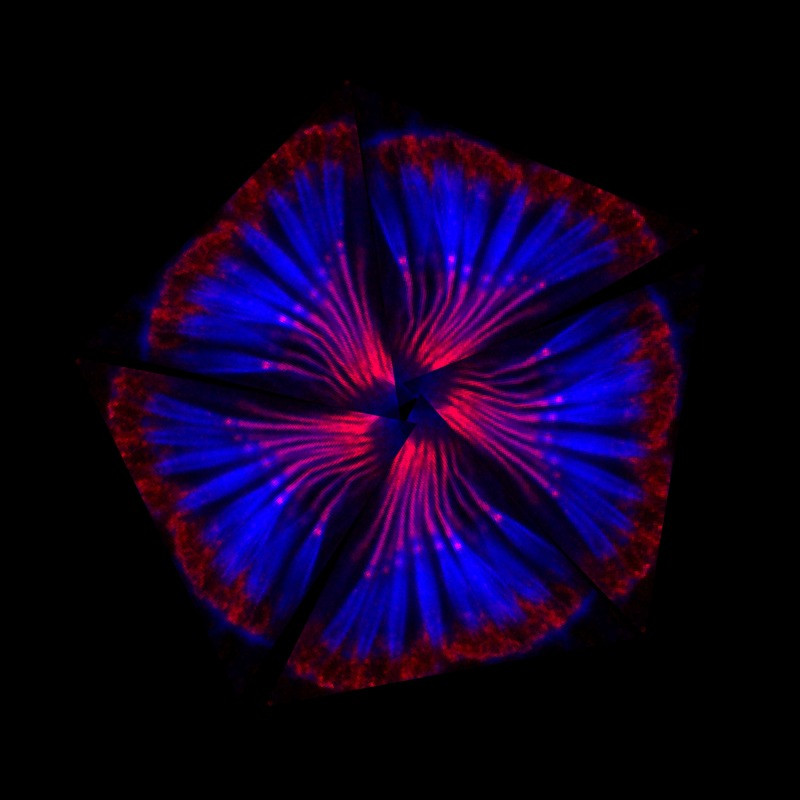 A montage of scolopidia – the structures that fruit flies require for hearing
A montage of scolopidia – the structures that fruit flies require for hearing
How Flies “Hear” Through Their Antennae
Instead of ears, flies utilize their antennae to detect sound. Despite the different anatomical structures, the underlying principles of hearing in flies and mammals share striking parallels. Within the second segment of a fly’s antenna are sensory structures called scolopidia. These scolopidia are suspended in the antenna and are responsible for sensing sound vibrations originating from the third antennal segment.
Similar to mammalian hair cells, fly scolopidia contain MET channels. Specialized cells called cap cells and scolopale cells anchor the tip of the scolopidia to the joint connecting the second and third antennal segments. This intricate structure allows sound waves to be transmitted to sensory neurons, activating the MET channels and generating electrical signals that the fly’s “brain” can process as sound.
Just as myosin motor proteins are crucial for mammalian hearing, they also play a vital role in the auditory system of Drosophila. Research has shown that myosin VIIa is essential for anchoring scolopidia to the antennal joint and ensuring the correct formation of the dendritic cap, a structure connecting sensory neurons to the joint.
Uncovering the Role of Ubr3: A Key Regulator in Hearing
Recent research published in eLife by Li et al. has identified a critical enzyme called Ubr3 that plays a regulatory role in both fly and mammalian hearing. This enzyme, an E3 ubiquitin ligase, was found to have a similar effect to inactivating myosin VIIa in flies. Specifically, the absence of Ubr3 causes scolopidia to detach and disrupts the normal distribution of proteins in the dendritic cap, mirroring the effects of myosin VIIa gene inactivation.
Further investigation revealed that Ubr3, along with another E3 ubiquitin ligase called Cul1, negatively regulates the addition of ubiquitin to myosin II. Ubiquitination is a process that can modify protein interactions and functions. In this context, Ubr3 and Cul1 help to control the interaction between myosin II and myosin VIIa. Proper regulation of this interaction is crucial for the correct localization of both myosin proteins and the fly equivalents of Usher proteins to the tip of the scolopidia. This localization is essential for the structural integrity and function of the scolopidia, and therefore, for the fly’s ability to hear.
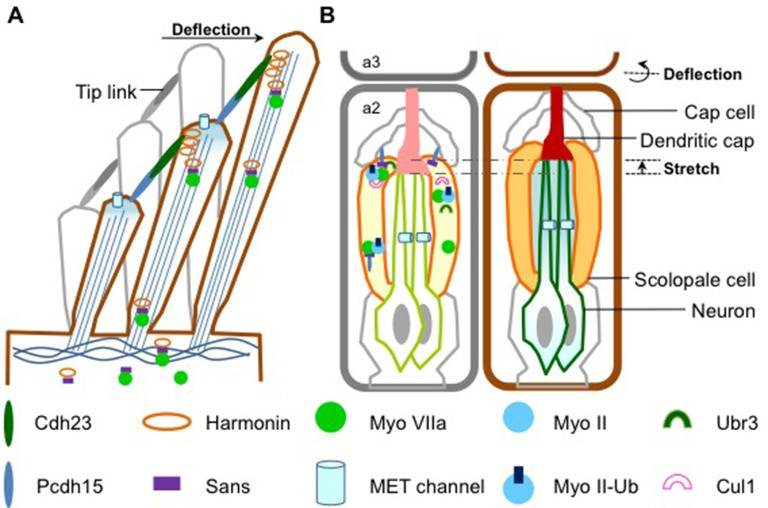 Figure illustrating sound detection in mammals and Drosophila antennae
Figure illustrating sound detection in mammals and Drosophila antennae
Figure 1: Comparative mechanisms of sound detection. (A) Mammalian hair cell stereocilia and MET channels. Myosin VIIa transports USH1 proteins for structural integrity. (Adapted from Richardson et al., 2011). (B) Drosophila antenna segment 2 with scolopidia and MET channels. Myosin II and VIIa, USH1 proteins, Ubr3, and Cul1 form a complex at the scolopale cell tip. Sound wave deflection opens MET channels. (Adapted from Boekhoff-Falk and Eberl, 2014).
Conserved Hearing Mechanisms: From Flies to Humans
The findings of Li et al. and their colleagues provide compelling evidence for a conserved molecular mechanism underlying hearing across species, from insects to mammals. This model highlights the importance of the Ubr3-mediated regulation of myosin II ubiquitination in promoting the formation of a myosin IIa-myosin VIIa complex (or its fly equivalent). This complex then facilitates the transport of USH1 proteins to the sound-sensing structures – stereocilia in mammals and scolopidia in flies – where they are essential for the function of MET channels and ultimately, for hearing.
By leveraging the powerful genetic tools available in Drosophila, researchers are continuing to identify new components and regulatory mechanisms involved in auditory organ development and function. These discoveries are not only expanding our understanding of insect sensory biology but are also providing valuable insights into the genes and proteins implicated in human deafness, paving the way for future therapeutic strategies for hearing disorders. So, the next time you try to swat a fly, remember they can hear you – and their tiny ears are revealing big secrets about hearing for all of us.
References
Li T, Giagtzoglou N, Eberl D, Nagarkar-Jaiswal S, Cai T, Godt D, Groves AK, Bellen HJ. 2016. The E3 ligase Ubr3 regulates Usher syndrome and MYH9 disorder proteins in the auditory organs of Drosophila and mammals. eLife 5:e15258. doi: 10.7554/eLife.15258
Boekhoff-Falk G, Eberl DF. 2014. Tectorial membrane and inner ear fluid mimicry in insect hearing. Developmental Neurobiology 74:82-97.
Kazmierczak P, Sakaguchi H, Tokita J, Wilson-Kubalek EM, Milligan RA, Muller U. 2007. Cadherin 23 and protocadherin 15 interact to form tip-link filaments in sensory hair cells. Nature 449:87-91.
Pan B, Zhang M. 2012. Mechanoelectrical transduction channel adaptation kinetics shape auditory receptor potential waveform. The Journal of Neuroscience 32:1189-1198.
Prosser HM, Davies KE, Glover L. 2008. Usher syndrome type I proteins: roles in neuronal development and function. Human Molecular Genetics 17:R201-R210.
Richardson GP, Gillespie PG, Holme RH. 2011. Usher syndrome: mechanisms of hair cell dysfunction and implications for therapy. Annu Rev Neurosci 34:209-229.
Todi SV, Charng WL, Pushkin V, Setsuko C, Marek KW,ிறோம் Eberl DF. 2005. Myosin VIIA is required for structural integrity of Drosophila auditory organs and development of auditory function. The Journal of Neuroscience 25:1119-1128.
Todi SV, Vardarajan V, Charng WL, Knowles JA, Marek KW, Eberl DF. 2008. The role of Drosophila MyoVIIA in the localization of the transmembrane protein Crumbs in photoreceptor and auditory organs. Molecular Biology of the Cell 19:3815-3827.
Weil D, Blanchard S, Kaplan J, Guilford P, Gibson F, Walsh J, Mburu P, Varela A, Levilliers J, Weston MD, et al. 1995. Defective myosin VIIA gene responsible for Usher syndrome type 1B. Nature 374:60-61.